Analysis of a DNA sequence can be accomplished via a method called electrophoresis. Electrophoresis is a term that basically describes the movement of molecules by way of an electric current and separation of those molecules based on size. This process occurs through agarose or polyacrylamide gels, which serve as a way to limit migration of molecules as they move from the negative anode to positive anode. Small molecules move through the gel matrix faster than larger molecules.
How does it work?
Each phosphate group from a DNA molecule is ionized
↓
DNA becomes negatively charged
↓
DNA migrates towards the positive pole (anode)
Factors Affecting Electrophoretic Separation
Strength of the electric current (voltage)
Concentration and type of the buffer
Gel density
Size of the DNA
AGAROSE CONCENTRATION AND SEPARATION RANGES
Agarose Concentration (%)
Separation Range (base pair size)
0.3
5,000 – 60,000
0.6
1,000 – 20,000
0.8
800 – 10,000
1.0
400 – 8,000
1.2
300 – 7,000
1.5
200 – 4,000
2.0
100 – 3,000
As agarose concentration increases, the separation range decreases
TYPES OF ELECTROPHORESIS SYSTEMS
Pulsed Field Electrophoresis
Best for very large DNA molecules
Current is applied in alternating directions
Field Inversion Gel Electrophoresis
(FIGE)
Alternates the + and – electrodes
Requires temperature controls and a switching mechanism
Polyacrylamide Gel Electrophoresis
(PAGE)
Best for very small DNA fragments
Initially used for protein separation, but can also be used for high resolution of nucleic acids
Capillary Electrophoresis
Molecules are separated by size and charge
Small Molecules = Fast
Large Molecules = Slow
Negatively Charged = Fast
Positively Charged = Slow
Utilizes a polymer inside of a capillary instead of a gel
Increased sensitivity
Understanding Buffer Systems
In order to change the pH of a buffered solution by one point, either the acidic or basic form of the buffer must be brought to a concentration 1/10th that of the other form.
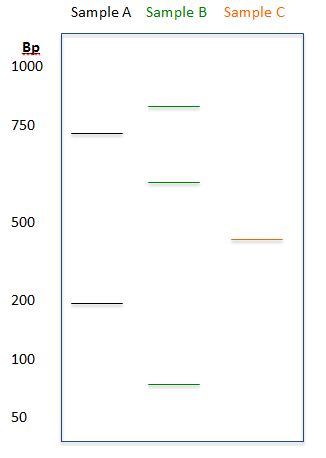
Test Your Knowledge
Based on the diagram, determine the sizes (to the best approximation) of the DNA fragments for each of the samples:
Answer:
Sample A: 200bp, 700bp
Sample B: 90bp, 600bp, 875bp
Sample C: 400bp
-LeAnne Noll, BS, MB(ASCP)CM is a molecular technologist in Wisconsin and was recognized as one of ASCP’s Top Five from the 40 Under Forty Program in 2015.
If you’re involved with molecular diagnostics, then L. J. Lee’s team at the Ohio State University would like your input. They’ve developed a new technology using molecular beacons and would love for lab directors, managers, and bench technologists to answer this short survey.
When considering methods to extract nucleic acids (DNA and RNA) to be used in downstream applications, it is important to think about what the sample source will be, as well as if it will be able to be used in an eventual assay. Being able to extract and purify DNA can lead to important information on infectious disease testing as well as patient genotype analysis. Purification of RNA can aid in understanding gene expression. However, once nucleic acid is extracted from a specimen, an additional set of parameters and guidelines are put in place to ensure that quality standards are met and the nucleic acid has not been compromised, ultimately leading to its ability to be used in patient testing.
Specimens / Sample Sources
Fresh Tissue in the form of cells such as, buccal cells, swabbed cervical cells, and even fresh DNA from the root of a single hair are all easily collected, stored, and provide enough DNA to be used in downstream PCR assays.
Whole Blood samples are easily available and require relatively simple methods to extract nucleic acid. However, the main source of issues comes from the type of anticoagulant that the blood specimen is mixed with. EDTA, Heparin, and ACD are all available as coating in blood collection tubes and are used to prevent clotting of the sample. Studies have shown that EDTA and ACD are acceptable anticoagulants that minimally interfere the quality and quantity of extracted nucleic acid as well as downstream PCR applications. Conversely, heparin is not completely removed during extraction procedures and can have an inhibitory effect on PCR and other enzyme based assays. Methods to eliminate heparin from a sample can be employed and include ethanol precipitation, boiling, and filtering.
Formalin-Fixed, Paraffin-Embedded Tissues provide a wealth of samples as these types of preserved tissues are typically stored in great numbers by pathology departments. The problem with tissues preserved in this manner is that quality is definitely compromised. When nucleic acids are exposed to fixatives, such as formalin, they become seriously fragmented and degraded. While these sources of samples aren’t ideal, there still are many benefits to using formalin-fixed or paraffin-embedded samples.
Extraction Methods
While most high volume laboratories employ automated extraction methods carried out on large pieces of instrumentation, there are laboratories that perform manual extractions at the bench. Manual extraction provides some of the best quality of nucleic acid samples. Typically, the purity of the nucleic acid is slightly compromised when automated methodology is employed; however the effect is somewhat negligible and allows a high volume of samples to be processed in various downstream applications.
Regardless of the sample type, the first steps of any extraction involve tissue isolation, disruption, and lysis of the cells. Paraffin-embedded samples require an initial step to remove the paraffin from the sample and this is accomplished via heat or the use of xylene. When isolating nucleic acids from blood, the white blood cells are isolated prior to extraction. Molecular detergents, such as SDS (Sodium Dodecyl Sulfate) are utilized to lyse the cells. Sometimes laboratories include a Proteinase K step due to the high amount of protein present in cell lysates.
Extraction methods vary, but the most common fall into three typical groups: Organic, Inorganic, or Solid Phase. Organic extractions utilize organic chemicals such as phenol or chloroform. Inorganic extractions use inorganic chemicals such as detergents, EDTA, acetic acid, or salt. Solid phase extractions immobilize nucleic acid on solid support system such as a spin columns or beads. Many automated extraction instruments employ silica bead based extraction chemistry.
Quality Assessment of DNA (Measurements)
Many PCR assays require a quality check for the nucleic acid purity and/or concentration. Spectrophotometers are an easy way to assess these two quality measures. Nucleic acids exhibit maximum absorption at 260nm and proteins at 280nm. Therefore, purity of a sample is assessed by measuring the optical density (OD) at 260nm and 280nm and calculating the ratio: OD260/280. A nucleic acid purity ratio of 1.8 – 2.0 is considered relatively pure. A reading less than 1.8 suggests protein contamination and readings above 2.0 suggest increased presence of RNA.
Note: An OD260 of 1.0 corresponds to 50 μg/mL of double stranded DNA or 40 μg/mL of RNA
A simple formula is used to calculate the quantity (concentration) of nucleic acid:
A= εbc
Where: A = Absorbance
ε = molar absorptivity
DNA is 50 L/mol-cm and RNA is 40 L/mol-cm
b = Path length (cm)
c = Concentration (mg/L)
Storage of Nucleic Acid
DNA can safely be stored long term if stored in Tris-EDTA buffer at 4°C. Typically, the colder the temperature, the less chance for degradation. Ideally, store DNA at -80°C and reduce the amount of freeze-thaw cycles. RNA should be stored in the same type of buffer at -80°C.
Test your Knowledge!
The following DNA samples are extracted from a whole blood sample and assessed on a spectrophotometer. Calculate the purity of each sample and comment on its quality:
Sample
A260
A280
1
0.500
0.270
2
0.320
0.310
3
0.445
0.219
Calculate the concentration of DNA and RNA if the A260 reading = 0.225 at a 1:100 dilution and the spectrophotometer has a 1.0 cm pathlength.
Answers
1.
Sample 1 = 1.85. The purity is considered ideal for DNA
Sample 2 = 1.00 which suggests protein contamination
Sample 3 = 2.11 which is ok for RNA
2. Solving for concentration (“c” from the formula above)
DNA: 1125 μg/mL
RNA: 900 μg/mL
-LeAnne Noll, BS, MB(ASCP)CM is a molecular technologist at Children’s Hospital of Wisconsin and was recognized as one of ASCP’s Top Five from the 40 Under Forty Program in 2015.
Dr. Diamond from the Washington University School of Medicine talked with Lab Medicine about all things Zika Virus: a brief history of the virus, modes of transmission, and the implications for laboratory professionals and pathologists.
One of the first things students of biology learn is basic molecular theory or, the central dogma of biology. The central dogma of biology states that “DNA makes RNA and RNA makes protein.” We know that the process of converting DNA to protein is not one involving magic, but rather more intricate methods termed, transcription and translation. For a quick review, check out my post A Primer on DNA and the Consequences of Mutation.
As we take a closer look at transcription, we will investigate how DNA is converted into pre-messenger RNA and ultimately into messenger RNA. The manner in which this occurs is termed RNA Splicing. Following splicing, mRNA is transported out of the nucleus to indirectly assist with protein synthesis.
In most eukaryotic genes, coding regions are interrupted by non-coding regions. Coding regions are termed exons, where alternatively non-coding regions are called introns. The coding region made up of exons is also called the open reading frame. During the process of transcription, DNA of an entire gene is translated into RNA, more specifically termed pre-messenger RNA (pre-mRNA). Pre-mRNA includes both exons and introns. Through a process, called RNA splicing, the introns of the gene are removed allowing the exons to be joined forming a continuous coding sequence. The continuous coding sequence is referred to as messenger RNA. mRNA contains a 5’ cap consisting of 7 methyl guanosine, a 3’ polyA tail, and is ready for translation into protein.
EXONS–Coding regions of the DNA that make up the mature messenger RNA
INTRONS–Non-coding regions that are removed from the DNA sequence to form mature messenger RNA
PROMOTER–Found at the 5′ end of the DNA sequence and functions to initiate transcription
Mature mRNA: 5′ 7 Me G–111122222222222222233333–AAAAA 3′
Test your Knowledge!
What is the function of the 5’ 7-methyl guanosine cap?
What enzyme is responsible for the addition of the polyA tail to the 3’ end of the mRNA?
What structures of the mRNA are translated into protein?
Answers
The 7-methyl guanosine cap is recognized as a site that signals the start of translation
Polyadenylate polymerase adds the multiple adenines to the 3’ end of an mRNA sequence
Introns are spliced out of the open reading frame and the exons are translated into protein
-LeAnne Noll, BS, MB(ASCP)CM is a molecular technologist at Children’s Hospital of Wisconsin and was recognized as one of ASCP’s Top Five from the 40 Under Forty Program in 2015.
In a post back in September, I quickly summarized the abnormalities that can occur with chromosomes as a whole (such as deletions, insertions, transversions, etc). There is so much more to learn (more than I could possibly put into one blog post), because the way chromosomes behave, depends on their structure and DNA sequence. For instance, genes with the same DNA sequence will behave differently depending on where they are located on a chromosome as well as the effect of the surrounding DNA sequence.
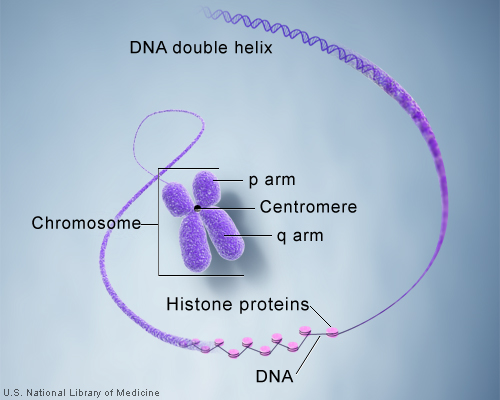
So how exactly is the immense length of DNA compacted into a chromosome? Let’s take a DNA sequence and see just how it makes up a chromosome. A single molecule of DNA spools around histone protein cores forming bead like structures called nucleosomes. Between each nucleosome is a sequence of DNA termed “linker DNA.” The amino acids associated with histones are lysine and arginine. The super coiled form is compacted and can be visualized as a karyotype in laboratory testing.
The centromere is the connection point of the duplicated chromosome, while telomeres are the endpoints. The short arm of the chromosome is termed “p” and the long arm of the chromosome is termed “q.” If we take these two chromosome arms into consideration, there are three types of chromosome morphology:
Metacentric – Chromosome arms are equal in length
Sub-centric – One arm is longer than the other
Acro-centric / Telocentric – One arm is extremely small or even missing
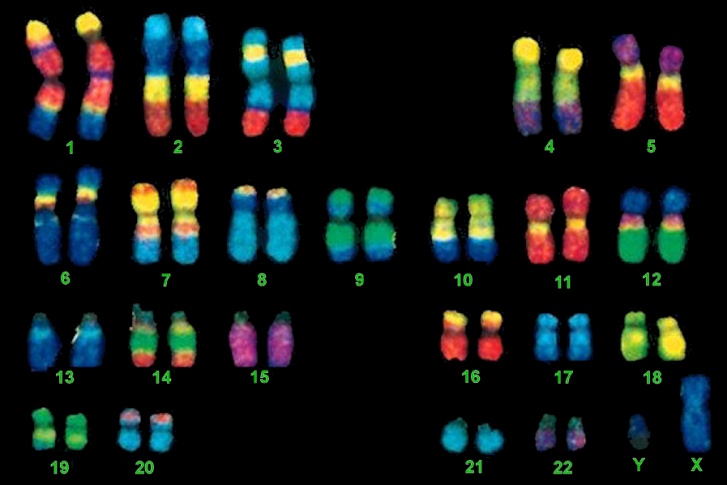
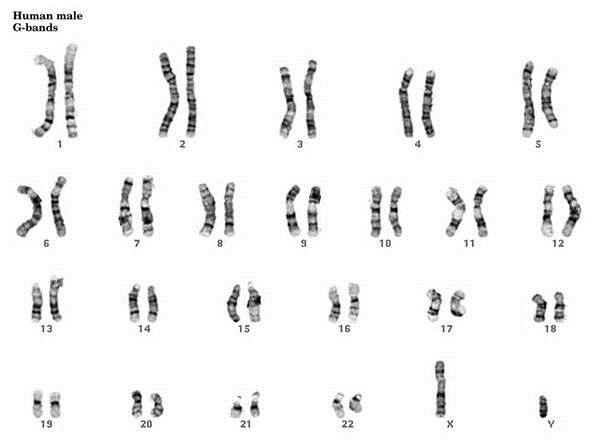
Chromosomal Staining Methods
As I mentioned above the complete set of chromosomes for an individual can be visualized via a karyotype. I’ve listed a few of the ways this can be accomplished:
G-Banding – Chromosomes are stained with giesma stains. The appearance differs based on the treatment of chromosomes prior to staining.
Q-Banding – Chromosomes are stained with fluorescent dyes, quinacrine or quinacrine mustard. Q-Band staining is similar to G-banding in that the fluorescent regions represent the AT-rich regions of the chromosome.
R-Banding – Results from heat treatment in a phosphate buffer followed by staining with Giesma dyes.
C-Banding – Centromere staining that results from alkali treatment.
TYPE OF BANDING
STAINING SUMMARY
G-Banding
· Geisma stain
· AT-rich regions stain darker than GC-rich regions
Q-Banding
· Quinacrine fluorescent dye stains AT-rich regions
R-Banding
· Banding pattern is opposite G-banding
C-Banding
· Stains heterochromatic regions close to the centromeres
· Usually stains the entire long arm of the Y chromosome
So how do you exactly identify chromosomal location based on banding patterns?
In studying disease and mutation, we follow a specific type of nomenclature to designate the regions that are of interest to us. Let’s take for instance something like the 22q11.2 deletion. What do all of these numbers and letters mean? To quickly summarize, 22q11.2 deletion syndrome occurs from the deletion of a small piece of chromosome 22 at a location: q11.2
22q11.2 DELETION
22
· Chromosome 22
q
· Long arm of chromosome (q)
1
· Region 1
1
· Band 1
2
· Sub band 2
So now, when we add in karyotope information you might see something like the following:
46, XY, del(8)(q21)
When you break it down, it states the patient is male (XY) and has a deletion in the long arm (q) of chromosome 8 at region 2, band 1
Translocation nomenclature can get a little more confusing:
46, XX, t(3;12)(p12.1;p11)
This designates a female has a translocation between the short arms (p) of chromosomes 3 and 12 and region 1, band 2, sub band 1; and region 1 band 1 respectively.
An example of Down syndrome: 47, XX + 21 (Female has an extra chromosome 21)
An example of Klinefelter Syndrome: 47, XXY (Male with extra X chromosome)
What do “FISH” have to do with Molecular Biology?
FISH, an acronym for Fluorescent In-Situ Hybridization, is a method used to detect and visualize protein, RNA, and DNA structures in the cell. FISH analysis is a relatively fast method that provides great resolution as it incorporates fluorescent probes labeled for detection of specific regions, deletions, and translocations. The images below show the difference between FISH and Karyotype images.
Robertsonian translocations are of importance because they involve translocating most of one entire chromosome to the centromere of another chromosome. They can be balanced or unbalanced. A balanced translocation usually results in no health difficulties because there isn’t a gain or loss of genetic material. However, due to the duplication or deletion of genetic material in an unbalanced translocation, syndromes and other malformations are likely to occur. The chromosome pairs common for Robertsonian translocations include translocations between 13 and 14, 14 and 21, and 14 and 15.
During a Robertsonian Translocation two chromosomes (typically acrocentric in formation) will break apart at their centromeres. The long arms will fuse to form a single chromosome and the short arms will also join to form a product. Typically the product created by the short arms contains nonessential genes and is eventually lost through cell division. Most people with ROB have only 45 chromosomes in each cell containing all of the essential genetic material and appear normal.
An example of a balanced Robertsonian Translocation would be when the long arms of chromosomes 14 and 21 fuse together. Phenotypically, the heterozygous carrier would appear normal because there are two copies of the major chromosome arms, resulting in two copies of the essential genes. However, children of the carrier could inherit an unbalanced translocation that causes Trisomy 21 (Down Syndrome).
Test your Knowledge!
Which two amino acids are associated with histones?
What type of chromosome morphology is shown:
If you wanted to stain chromosomes to see varying regions that were AT-rich, which type of stain would you use?
Describe the following karyotype results: 46, XX, t(1;14)(p21.3; p17.6)
Answers
Lysine and Arginine
Sub-centric
G-Banding (although Q-Banding also will produce darker regions that are AT-rich)
Female patient, with a translocation between the p and q arms of chromosomes 1 and 14, and region 2, band 1, sub band 3; and region 1 band 7, sub band 6 respectively.
References:
Buckingham, L. (2012). Molecular Diagnostics: Fundamentals, Methods and Clinical Applications (2nd ed.). Philadelphia: F.A. Davis Company.
Coleman, W.B, Tsonagalis, G.J. (2005). Molecular Diagnostics: For the Clinical Laboratorian. New York: Springer-Verlang
Searle, B. Rarechromo.org. The Rare Chromosome Disorder Support Group. 1996. Web. 19 Dec. 2015.
-LeAnne Noll, BS, MB(ASCP)CM is a molecular technologist at Children’s Hospital of Wisconsin and was recognized as one of ASCP’s Top Five from the 40 Under Forty Program in 2015.
To gain a solid understanding of Molecular Diagnostics, one has to grasp the fundamentals of DNA Replication. The double helix nature of DNA requires numerous moving parts working together to produce two identical strands of DNA from one original DNA molecule.
The semi-conservative process of DNA replication occurs in a 5’ to 3’ anti-parallel direction. The replication process is described as semi-conservative because the sequence of nucleotides is maintained through new generations of replication. An extremely important enzyme involved in the beginning stages of DNA replication, is called Topoisomerase. It is responsible for regulating the over-winding and under-winding of DNA just ahead of the replication fork. Topoisomerase binds to the DNA then “cuts” the phosphate backbone so that the DNA can be unwound then resealed at the end of replication. Also, before replication can begin, an enzyme called helicase must first unwind and untangle the double-stranded DNA. Single stranded binding proteins (ssbp) prevent premature binding as well as protect the single stranded DNA from being digested by nucleases.
Leading Strand vs. Lagging Strand
During replication, two separate strands of DNA are formed in different ways. The lagging strand exhibits discontinuous 3’ to 5’ growth away from the replication fork and requires primase to “prime” the synthesis of the lagging strand. An RNA primer is added to the lagging strand of the DNA by RNA polymerase. This RNA primer begins synthesis of the lagging strand. The separate fragments of the lagging strand are termed Okazaki fragments. It’s important to note that due to the discontinuous formation of the lagging strand, each Okazaki fragment requires its own, separate, RNA primer. Finally, DNA ligase forms phophodiester bonds between the existing DNA strands to join the Okazaki fragments together. Alternatively, the leading strand during replication grows towards the replication fork in a 5’ to 3’ direction. The leading strand only needs one single RNA primer to immediately begin replication and therefore does not require DNA ligase.
LEADING STRAND SYNTHESIS
REQUIREMENTS
Toward Replication Fork
Single RNA Primer
5’→ 3’
Continuous Growth
LAGGING STRAND SYNTHESIS
REQUIREMENTS
Away from Replication Fork
Primase
3’→ 5’
Multiple RNA Primers
Discontinuous Growth
DNA Ligase
Creation of Okazaki Fragments
DNA Polymerase III and its Role in Replication
While you should become familiar with the extensive list of DNA Polymerases (shown below), the core polymerase involved in DNA replication is DNA Polymerase III. It functions as a catalyst in the formation of the phosphodiester bonds between an incoming deoxyribose nucleotide triphosphate (dNTP) determined by hydrogen bonding to the template at the 3’ end of the primer.
PROKARYOTIC DNA POLYMERASES
FUNCTION
DNA Polymerase I
Recombination, Repair, Replication
DNA Polymerase II
Repair
DNA Polymerase III
Core Polymerase
Replication
DNA Polymerase IV and V
Bypass DNA Damage (Y-Family DNA Polymerases)
EUKARYOTIC DNA POLYMERASES
FUNCTION
Alpha (α)
RNA Primase
Lagging Strand
Replication (Initiation, Okazaki Fragment Priming)
Beta (β)
DNA Repair
Delta (δ)
Leading Strand
Repair
Epsilon (ε)
Sensor of DNA replication that coordinates transcription cycle
Repair
Gamma (γ)
Mitochondrial Replication
RNA POLYMERASES
FUNCTION
RNA Polymerase I
rRNA (ribosomal RNA)
RNA Polymerase II
mRNA (messenger RNA)
RNA Polymerase III
tRNA (transfer RNA)
sbRNA (small nuclear RNA)
-LeAnne Noll, BS, MB(ASCP)CM is a molecular technologist at Children’s Hospital of Wisconsin and was recognized as one of ASCP’s Top Five from the 40 Under Forty Program in 2015.
Recently, I came across a research article that suggests an elevated germ-line mutation rate in teenage fathers, thus leading to an unexpectedly high level of DNA mutations in the children born to teenage fathers.
Many studies have been conducted on the theory that male germ cells go through a higher number of cell divisions when compared to that of female germ cells, and that the higher number of paternal cell divisions leads to an increased DNA mutation rate. The paper suggests that the increased presence of DNA mutations in sperm cells of teenage boys could explain why their offspring might be at higher risk for a spectrum of disorders when compared to parents in their twenties.
-LeAnne Noll, BS, MB(ASCP)CM is a molecular technologist at Children’s Hospital of Wisconsin and was recognized as one of ASCP’s Top Five from the 40 Under Forty Program in 2015.
We can’t talk about Molecular Diagnostics without possibly talking about DNA, and with DNA, comes mutation. Much of the work I do revolves around searching for specific variants in a patient’s DNA sequence. We all have mutations in our DNA, but does that mean we all are affected negatively? Absolutely not. Spontaneous mutations occur during normal process such as DNA replication and repair within our cells. They also can arise from exposure to ionizing radiation, UV exposure, and chemical agents. Some mutations are passed down through reproductive cells. Mutations can be categorized as harmless or sometimes hurtful giving rise to gene defects, copy number variants, metabolic deficiencies, and cancer, while others result in positive effects.
Where do mutations occur?
Somatic mutations occur in cells that aren’t reproductive in nature. These mutations for the most part do not have a blatant effect on the organism because our normal body cells are able to counteract the mutated cells. However, sometimes mutations in somatic cells can affect division of cells, which has the potential to result in forms of cancer. On the contrary, germ-line mutations occur in reproductive cells and have the possibility of being passed down through generations, resulting in the presence of the mutation in all of the organism’s cells.
Understanding DNA
A T G C bases, along with a sugar, and a phosphate group combine to form a polymer of nucleotides. This polymer backbone of alternating sugar and phosphates is what we call DNA (Deoxyribonucleic Acid). DNA is transcribed to RNA (Ribonucleic Acid) which instead of Thymine (T) contains a Uracil (U). Finally, RNA is translated to protein.
Pyrimidines
Purines
Single-ringed organic bases
Double-ringed organic bases
Thymine (T)
Cytosine (C)
Uracil (U)
Adenine (A)
Guanine (G)
Hydrogen Bonds Between Complimentary Strands of DNA
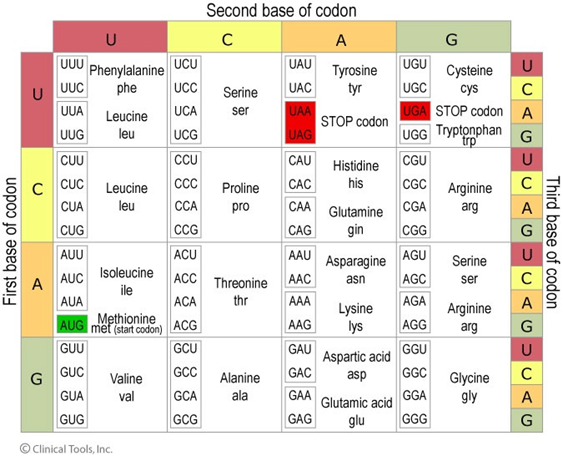
Amino acids are the triplet codons that make up proteins. It is extremely important to become familiar with the codon table. You don’t need to memorize it, however you should at a minimum know what triplets code for START and STOP codons. You will quickly see that there are multiple codes for a single amino acid. This becomes important when considering the effects of mutation:
Like I mentioned earlier, many of the tests I perform involve screening for variants in DNA sequence. I accomplish this through different methods, which I will touch on in future blog posts. We could easily discuss mutations for hours and hours, but for now, it’s probably most important to gain a quick (and simple) understanding of the types of mutations and what the resulting effects can be. For the sake of demonstration, let us consider the following normal (wild type) sequence pattern:
NOTE: Uracil replaces Thymine because of transcription from DNA to single stranded messenger RNA (mRNA)
Transition Mutations occur when mispairing results in a purine being replaced with a different purine, or a pyrimidine replaced with a different pyrimidine:
A→G
C→T
Transversion Mutations occur when a purine is replaced with a pyrimidine or vice versa:
A→T
C→G
Missense Mutations have the ability to result in a change in the amino acid. This happens when the codon triplet is altered, and the amino acid sequence of the encoded protein is changed:
Sometimes, Missense Mutations are silent:
Notice how the structure of the gene product is unchanged. This happens because all amino acids can be encoded by more than one triplet codon.
Frame shift mutations can result from either a single base insertion or a single base deletion.
Insertion.Deletion.
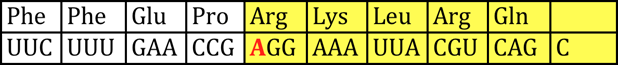
Nonsense mutations are exactly what they are called… Nonsense! Here, a base substitution changes an amino acid into a stop codon. Nonsense mutations result in abnormal termination of translation and the protein product is shortened:
Chromosomal Abnormalities
Sometimes changes in the actual chromosomal structure or number take place in the cells. These changes can result in deletions, duplications, inversions, and translocations of chromosomes. Chromosomal deletions and insertions are simply the loss or gain of chromosomal material. Inversions result from the removal, flipping, and then reconnection of the chromosomal material within the same chromosome.
Translocations are more complicated and involve switching of genetic material between two chromosomes. A reciprocal translocation occurs when parts from two different chromosomes exchange. We call a reciprocal translocation “balanced” when there isn’t a gain or loss of genetic material between the chromosomes (both chromosomes remain fully functional). An “unbalanced” reciprocal translocation affects an offspring’s phenotype due to extra or missing genes.
This information I have touched on is very general, however, it doesn’t mean it is not of importance. These basic concepts are the fundamentals needed to begin understanding mutation as they will provide us with the framework for recognizing the needs and importance of molecular diagnostic testing!
Test your Molecular Knowledge
What amino acids do the following codons code for:
GAU
AUG
UAG
CAG
UGA
Classify the following mutations:
T→A
T→C
AGA→UGA
AGA→AAA
AGA→CGA
Answers
1 a) Aspartic Acid b) Methionine (start) c) Stop codon d) Glutamine e) Stop codon. 2 a)Transversion b) Transition c) Nonsense d) Missense e) Silent
-LeAnne Noll, BS, MB(ASCP)CM is a molecular technologist at Children’s Hospital of Wisconsin and was recognized as one of ASCP’s Top Five from the 40 Under Forty Program in 2015.
While I have been working in the field of all things “Molecular” for close to twelve years, it wasn’t until early this year that I decided to actually sit for the ASCP certification exam in Molecular Biology. With over five years working in research and currently going on seven years in a clinical lab, I was feeling pretty confident about my knowledge and background, but then panic set in. Would I be able to pass a test? I haven’t taken an exam since my final years of college! I have two young children who are extremely active in extracurricular activities, a husband who also works full time. Needless to say, any extra time I had for myself would be spent studying. It didn’t take long to find out that while there was a very general outline of topics covered and an extensive list of textbooks to serve as a starting point, there were not specific details on where to focus my efforts. There was no single study guide, no tangible tools I could utilize to make exam preparation fit into my busy life any easier.
Fast forward to the present, where all of the panic was for not. In February I did pass (with flying colors) and happily sport MB (ASCP)CM after my name. When I was recently approached about writing on a regular basis for Lablogatory, I knew it would be an amazing opportunity to educate others on the exciting field of Molecular Diagnostics as well as combat the fear that comes with taking the Molecular Biology exam. I plan to focus on more specific areas in the field of Molecular Diagnostics that I came across in my studies, which will help others in their preparation for the MB(ASCP) exam. I will cover theory, applications, techniques, and practices. Also, keep an eye out for case study questions that I will provide to stretch your thinking through interpretation of molecular results, at the same time, keeping you up to date on hot topics in the field.
With all that said, the first most important suggestion I have is: spend time preparing to study. While it might seem silly to some, having a plan BEFORE you jump into study mode will actually set you up for success. All too often, just reading everything you can on every single topic will result in information overload and ultimately burn you out. You will quickly find yourself jumping all over the place frantically trying to memorize every detail you come across.
First, browse the content outline and choose some texts to review. Notice how I say, “Choose some?” Don’t feel like you have to read all of them. I went online and reviewed the texts, I asked colleagues for recommendations, then decided to purchase two books. You might find that you need more or less, just don’t go overboard.
Next organize a binder. I am extremely visual and hands on so I study best with things in front of me that aren’t electronic based. I divided my binder based on the content outline. You will come across papers, technical notes, and procedures that you will want to keep as study tools. Having a binder for all of these notes from multiple sources will keep you on track. It will also serve as a great reference guide for you as you move through your career.
Now is where you can get super retro. You may have noticed in my photos that I made flash cards! It is a little archaic and time consuming, but I knew I was going to be busy with my daughter’s traveling basketball team; therefore, I needed something small I could take with me to study while sitting on the bleachers. I am aware of software programs that allow you to make electronic based flash cards, so if that works better for you feel free to try that route.
My take home message is, obtain and prepare whatever tools you need to get yourself ready to study. Organization is key and setting aside time each day, even if only an hour to focus on one application or theory will definitely be to your benefit. Reach out to colleagues and gather as much information as you can. Once all of your tools are in place, it’s time to get to work! For anyone interested in obtaining their MB(ASCP) certification, I urge you to check out the content outline and list of texts on the ASCP Board of Certification site.
-LeAnne Noll, BS, MB(ASCP)CM is a molecular technologist at Children’s Hospital of Wisconsin and was recognized as one of ASCP’s Top Five from the 40 Under Forty Program in 2015.

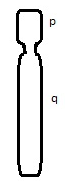









{kind=link}
{kind=link}